Infection – an adjuvant for autoimmunity: adjuvant effect
- EPA Announces First-Ever Regulation for “Forever Chemicals” in Drinking Water
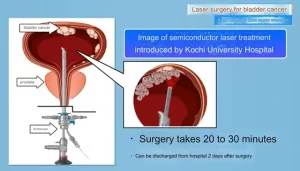
- Kochi University pioneers outpatient bladder cancer treatment using semiconductor lasers
- ASPEN 2024: Nutritional Therapy Strategies for Cancer and Critically Ill Patients
- Which lung cancer patients can benefit from neoadjuvant immunotherapy?
- Heme Iron Absorption: Why Meat Matters for Women’s Iron Needs
- “Miracle Weight-loss Drug” Semaglutide Is Not Always Effective
Infection – an adjuvant for autoimmunity: adjuvant effect
Infection – an adjuvant for autoimmunity: adjuvant effect. The history of the concept of host immunity can be traced back to the 5th century BC. The ancient Greek historian Thucydides discovered that when the survivors of the plague were exposed again to the environment that caused the plague, their condition did not worsen. Around the 10th century in China, it was also discovered that survivors of smallpox would be protected from re-infection of the disease. At the same time, in the practice of smallpox vaccination, people began to introduce smallpox (smallpox) scabs or pustules into the skin or nose of patients to prevent smallpox infection.
In 1796, Edward Jenner (1749-1823) invented vaccination, inoculating vaccinia virus before infection to prevent smallpox from spreading in humans. When the body is infected, the high immune response caused by protection may be harmful to the body. More than a century ago, the first case of human autoimmune disease-paroxysmal hemoglobinuria (Donat-Landsteiner syndrome) was considered to be a late symptom of syphilis. Studies have shown that rheumatic fever and rheumatic heart disease are caused by streptococcal infection, and viral myocarditis is related to type 1 diabetes and Coxsackie virus infection.
Sir William Osler (1849-1919) pointed out that the cause of death in some patients is the body’s immune response rather than infection. Human autoimmune diseases result from the poor inheritance of a group of genes that collectively control and increase autoimmune response. The most prominent susceptibility gene is a member of the major histocompatibility complex (MHC) encoded by the human leukocyte antigen gene. In addition to genetic susceptibility, environmental triggers (such as infection) are necessary in the onset of human autoimmune diseases.
According to the two signal hypotheses, viruses and bacteria can provide the autoantigen-specific signals required to activate T cell receptors (TCR). The uncontrollable immune response caused by endogenous autoantigens is the cause of autoimmune diseases. Several mechanisms have been proposed to explain the autoantigen-specific response after infection, including molecular simulation, T cell degeneration, epitope expansion, the release of usually invisible or concealed antigens, and changes in autoantigens.
Until recent years, less attention has been paid to non-clonal and non-specific second signals that can stimulate and worsen the autoimmune response. From a historical point of view, in the twentieth century, the specific recognition mechanism of immunoglobulin, TCR and clonal selection to identify pathogen antigens is dominant in immune research and theory. However, the survival of the infected host depends on the inherent structure of immune molecules that can adapt to the receptor. For example, the antigen presentation (signal 1) and the expression of costimulatory molecules (signal 2) by the innate immune antigen presenting cells (APC) (such as macrophages and dendritic cells) necessary for the activation of adaptive immunity.
Importantly, we have learned that “adjuvants” can greatly enhance antigen presentation in the body, including stimulation by mixtures developed through experience. In fact, the form of aluminum salt adjuvant vaccine has been widely used, but its mechanism of action is not fully understood. Charles Janway, JR (1943–2003) called adjuvants as the “dirty little secret” of immunologists. The new understanding of the pattern recognition of the innate immunity of pathogens helps explain the “auxiliary effect”.
Adjuvant effect
The adjuvant effect “is used for the first time in the key role of Mycobacterium tuberculosis in Freund’s induction of experimental thyroiditis. It refers to the use of an exogenous microbial factor that promotes an antigen-specific immune response in the body together with an antigen. These inherent properties The immune system allows the injected antigen to be more effectively discovered and captured by the adaptive immune system, which leads to the enhancement of the efficacy of T cells and clonal expansion.
Similarly, in terms of autoantigens, adjuvants are also responsible for activating the innate immune response to drive the development of autoimmune diseases. In other cases, the central and peripheral immune tolerance mechanisms for autoreactive T cells to escape are triggered by exogenous adjuvants to become autoimmune adjuvants. Adjuvants can activate APC to overcome the inhibitory effects of CD4 + and CD25 + regulatory T cells, leading to the activation of autoreactive T cells. Adjuvants are usually based on the composition of microorganisms and have strong biological activity. Infectious pathogens naturally produce their own adjuvant effects and can induce autoimmunity.
To explain this phenomenon, Janway proposed in 1989 that the evolutionarily conserved molecular components of infectious microorganisms-pathogen-associated molecular patterns (PAMPs)-encoded by a limited set of non-clonal germ lines are called ” Pattern recognition receptors (PRRs) are specialized receptors for the innate immune system to detect.
Supported by Janway’s hypothesis, Hoffman and his colleagues published a report on the Toll-mediated antifungal immune response in Drosophila in 1996. Nearly ten years ago, Nüsslein-Volhard and his assistants participated. Type I transmembrane protein was found in the experiment of dorsoventral polarity development of Drosophila embryos. In 1997, Janway and colleagues reported on an evolutionarily conserved human homolog of Drosophila Toll, now called TLR4, which is a human Toll-like receptor (TLR). TLR4 was found to contain an extracellular leucine-rich repeat (LRR) domain and an intracellular TIR domain.
It is currently believed that there are about 10 human TLRs as PRRSs that bind PAMPs (or co-receptors bind PAMPs) through their LRR-containing extracellular domain. The distinctive cytoplasmic TIR domain receptor intracellular signaling induces effective antibacterial and pro-inflammatory responses. The definition of PAMPS has been expanded, eliminating the need to identify each pathogen for structural evaluation, resulting in the concept of “microbial-related molecular patterns” (MAMPs).
Subsequent studies found that the release of an inflammatory response from endogenous molecules can also activate Toll-like receptors and lead to the use of “danger/damage-related molecular patterns” (DAMPs), largely based on endogenous host molecular signaling. The concept of harm or damage to the immune system. Innate immune response mediates adjuvant effects. PRRs recognize PAMPs/MAMPs and DAMPs, respectively, and are alert to the presence of infection and tissue damage in the immune system. PRR participates in the adjuvant effect of APC to provide the necessary costimulatory signal 2 and the production of inflammatory cytokines and to make antigen-specific adaptive immune response.
TLRs represent a family of single-span membrane PRRs located on the cell surface and endosomal membrane. In short, TLRs play a vital role in innate immunity against the immune response of potentially harmful microorganisms. In addition, it is also a biosensor for tissue damage. The ability of TLRs to regulate cellular responses to pathogens determines the corresponding immune response to infection. The combination of specific PAMPs and TLRs guides the combination of TLR linker molecules to activate a command that acts on the transactivation of nuclear reverse transcription factors.
The expression of inflammatory cytokines induced by activated NF-κB further enhances the inflammatory response. Activated interferon regulatory factor 3 (IRF-3) induces the transcription of irf-3-responsive genes. TLR and other innate immune PRRS play an important role in regulating adaptive immunity. In addition, TLRs are also effective mediators of sterile, non-infectious inflammatory diseases (atherosclerosis, myocardial ischemia-reperfusion injury, transplant rejection, and many autoimmune diseases).
The innate immune system can also sense the proteolytic enzymes produced during the infection process from the host and pathogens. When exposed to pathogens, host cells often encounter multiple PRR agonists at the same time. When activated by various microbial components, such as PAMPs and proteases, the coordinated interaction of signal transduction pathways can be triggered by innate immune PRRS, and regulate inflammation through synergy or antagonism. Therefore, the integration of signals from multiple PRRs may drive specific inflammatory responses to a combination of environmental stimuli.
Auxiliary autoimmunity of virus infection:
Evidence of viral myocarditis caused by Coxsackie virus About half of human infectious myocarditis was previously infected with an acute virus. Myocarditis is a serious heart disease, especially for young people. It can progress to dilated cardiomyopathy and heart failure requiring heart transplantation. It is usually caused by Coxsackievirus B3 virus (CB3) and a positive-strand RNA enterovirus of the picornaviridae family. Human infectious myocarditis can be reproduced in a mouse experimental model of myocarditis.
In the chronic phase, non-infectious viral RNA can be detected. However, the chronic phase of viral myocarditis is an experimental autoimmune myocarditis induced by immunity to cardiac myosin or its peptides. It must be pointed out that myocardial myosin immunity requires continuous stimulation of powerful CFA emulsions to induce disease.
If myocardial myosin is emulsified, cardiac inflammation will not occur, instead of being present in IFA. Therefore, the inflammatory cytokine environment produced by viral infection or exogenously effective adjuvant determines the degree of inflammation, and the degree of inflammation determines the rate of progression of chronic myocarditis. During infection, genetically determined viral elements are perceived as PAMPs through TLRs and other PRRs.
Viral RNA is recognized by Toll-like receptors 3, 7, and 8, which are also RNA helicase cell sensors. The TLR3 mutant encoding, enteroviral myocarditis, and dilated cardiomyopathy in patients indicate that the CB3-mediated IFN-β antiviral response failed to control CB3 replication in vitro. The structure-independent PAR2 inhibits the IFN-γ and β antiviral responses induced by TLR3, thereby aggravating the myocarditis induced by CB3 (Weithuser et al., 2013).
In contrast, the thrombin receptor PAR1 promotes the IFN-γ and β antiviral responses induced by TLR3, thereby protecting viral myocarditis caused by CB3. Effective antiviral immune responses generally have protective effects, and they contribute to the development of several autoimmune diseases in humans.
CB3 infection also activates other innate PRRS to induce the expression of pro-inflammatory cytokines (such as IL-1 and TNF-α), which is critical for the development of autoimmune myocarditis after viral infection. TLR4, TLR7 and TLR8 have been reported to mediate CB3-induced cytokine production in vitro, while TLR4 and TLR9 promote CB3-induced acute myocarditis in vivo. MDA5 and adaptor molecule protein (MAVS) are the key to survival during CB3 infection factor. In CB3 infection-susceptible mice, exogenous IL-1 aggravated myocarditis, and blocking IL-1 can significantly reduce the severity of the disease.
The production of the anti-inflammatory cytokine IL-10 protects myocarditis induced by CB3 or myosin CFA immunity. In mice against chronic myocarditis (such as B10), resistance can be overcome, and if IL-1, TNF-α, or induced LPS play a role during CB3 infection, the disease will be induced. IL-1 and TNF-α may increase local inflammation and cause tissue damage. These cytokines can also induce APCs to provide stimulation signals to activate non-antigen-specific T cells through adjuvant effects.
In the combined adjuvant of the EAM model, the mycobacterial extract and pertussis toxin partly show signals through TLR4. In addition, muramyl dipeptide components extracted from Mycobacterium tuberculosis also activate inflammasomes, which requires LPS/TLR4 to start. Therefore, the observed similarity in the development of CB3-induced myocarditis and the EAM model may be due to the ability of PAMPs in CB3 and the mycobacterial component of CFA to carry out the innate immune response process of PRRs. In the early stage of infection, CB3 replicates to inhibit the TLR4 defect center, which is consistent with the antiviral function in the tlr4-irf-3 pathway.
However, the immune phase after TLR4 activation promotes CB3 induced autoimmune diseases. This observation is consistent with a recent report that the absence of TLR4 in CD4 + T cells makes the autoimmune inflammatory effect almost disappear, and then may slow down the T helper 17 (Th17) response. Therefore, in the process of innate immune response, TLR4 activates the immune system to alert the host to prevent virus replication, but then may promote autoimmune diseases.
In addition, TLR4 also feels that endogenous injury signals further increase the local inflammatory response and contribute to the adjuvant effect. As “adjuvant diseases” are strong promoters of adjuvant immune response, the question has been raised whether adjuvants can be induced alone in human autoimmune diseases. The precedent for this possibility was included in the management of CFA by Pearson in 1956, rather than a report during the IFA period, which caused an inflammatory single or multiple arthritis in Wistar and Long-Evans rats.
Other systems include the eyes, skin, gastrointestinal tract, and urogenital mucosa. The mechanism of this disease is not yet clear, but there is a possibility that in these special animals, adjuvants have been carried out to enhance the autoimmune process. So far, humans have not reported similar results. Similar vaccines used in the United States have no adjuvants. The two different formulations of reported adverse reaction rates are comparable. Using adjuvant effects and autoimmune diseases to prevent cancer. When immune surveillance is weakened, its own disadvantage is that it will contribute to the development of cancer.
Non-clonal adjuvants can be used to achieve immune enhancement and improve anti-cancer immune response. Historically, severe infections in tumor-bearing patients led to the spontaneous regression of tumors. William Conley (1862–1936) pioneered the use of pollock toxins, including extracts that kill Gram-positive Streptococcus pyogenes and Gram-negative Serratia marcescens.
The preparation has been reported to be injected directly into the tumor site to successfully treat various tumors. Bladder perfusion BCG (BCG) – Guérin is an effective single-stranded RNA combined with TLR7, an agonist of imiquimod for the treatment of superficial bladder cancer. It has been successfully used as a skin cream for the treatment of tumors. Tumor cells have been genetically engineered to express bacterial flagellin, activate TLR5, and recently induce effective tumor-specific immune responses in the body.
Although there are several mechanisms that may be involved in the anti-tumor activity of these immunomodulators, the activation of the innate immune system through specific PRR may be the center of infection and/or tissue damage to drive the inflammatory response and autoimmune-driven adjuvant effects.
Conclusion:
In the twentieth century, adjuvant concepts based on observations and experiments and breakthroughs in immune theories played an important role in promoting our understanding of the immune system. Adjuvants and natural infections can exert effective immunostimulatory activity through adjuvant effects, which can lead to autoimmune diseases. The powerful adjuvant effect can also be used to develop new anti-tumor therapies and effective vaccines.
The goal of the vaccine is to reproduce the protection provided by the infection while minimizing the risk. The latest advances in the understanding of the molecular mechanisms involved in immune cell activation provide opportunities for fine-tuning the development of vaccines and provide strategies for adjuvant effects to identify receptors by activating specific immune patterns.
Disclaimer of medicaltrend.org